Abstract
Over the past 3 decades, the global emergence of Escherichia coli sequence type 131 (ST131) has posed a significant public health concern, making it one of the most notable multidrug-resistant pathogens. ST131 is responsible for urinary tract infections and bloodstream infections (bacteraemia) that affect people of all ages worldwide. First identified in 2008, it has rapidly spread across various continents, demonstrating its remarkable ability to disseminate. Its success is attributed to a combination of antimicrobial resistance gene acquisition, efficient host colonisation, and transmission dynamics. Although not hypervirulent, ST131 outcompetes commensal E. coli in gut colonisation and persists asymptomatically in carriers, facilitating community and hospital spread. Key to its dominance are mobile genetic elements (e.g., incompatibility group F plasmids carrying extended-spectrum β-lactamase blaCTX-M-15) and clonal sub-lineages, such as H30-Rx, which combine resistance and virulence traits. However, the relative contributions of antimicrobial resistance, metabolic adaptation, and host factors remain a topic of debate. This review synthesises evidence on the evolutionary, ecological, and epidemiological drivers of ST131’s dissemination, highlighting gaps in understanding its persistence and transmission. Addressing these gaps is critical for developing targeted interventions against this multidrug-resistant clone.
Key Points
1.Over the last 3 decades, Escherichia coli sequence type 131 has become a prominent multidrug-resistant pathogen, causing a significant number of urinary tract and bloodstream infections worldwide, and it has rapidly spread across continents since its discovery.2.Its success can be attributed to the acquisition of antimicrobial resistance genes, as well as its ability to efficiently colonise the gut, facilitate asymptomatic carriage, and transmit in both community and healthcare environments.
3.Although sequence type 131 is not classified as hypervirulent, it possesses advantages in resistance and colonisation; however, the specific contributions of antimicrobial resistance, metabolic changes, and host factors to its prevalence are still not fully understood, emphasising the necessity for focused research to guide control measures.
INTRODUCTION
Escherichia coli is a Gram-negative bacillus that constitutes a significant element of the human gut microbiome. Despite its normal presence in healthy individuals, it possesses pathogenic strains that can lead to a range of diseases, including diarrhoea, urinary tract infections (UTI), meningitis, and sepsis.1,2 The spread of multidrug-resistant (MDR) E. coli is a significant public health concern worldwide.3-5 Based on infection site, virulence factors, and clinical manifestations, Peirano et al.6 categorise E. coli pathotypes into: 1) normal gastrointestinal flora of humans and animals; 2) E. coli causing gastrointestinal infections; and 3) extraintestinal pathogenic E. coli (ExPEC), leading to infections outside the gastrointestinal tract. Among these, the pandemic ExPEC sequence type (ST) 131 has emerged as one of the most successful MDR strains.7 For instance, in North America, approximately 50% of extended-spectrum β-lactamase (ESBL) producing and 20% of fluoroquinolone-resistant E. coli are linked to ST131.8
Moreover, it affects adult and paediatric populations globally and causes UTIs and bloodstream infections.7,9,10 For example, these organisms are responsible for an estimated annual incidence of up to 7% of community-acquired UTIs in the USA and globally. Among these cases, post-menopausal women are at risk of developing severe and drug-resistant forms of UTI.7 Additionally, there has been a notable increase in the faecal carriage of ST131 within asymptomatic individuals.11,12 For instance, studies on the prevalence of E. coli ST131 among ESBL-producing or fluoroquinolone-resistant E. coli isolates colonising the digestive tracts found notably high rates among nursing home residents, reaching 44% in Germany (2010–2011) and 98% in the UK (2005–2006), as well as among children in French daycare centres, with a rate of 44%.3,13-14
ST131 was first reported in 200815-17 and spread to nine countries across three continents, namely North America (Canada), Europe (France, Portugal, Spain, Switzerland), and Asia (India, South Korea, Kuwait, and Lebanon), by 2009.3,17,18 Recent research indicates that the prevalence of ST131 among human clinical E. coli isolates varies by geographic location and host group (overall rates ranging from 12.5% to nearly 30%).17,19-21 The emergence of these pathogens is linked to the acquisition of ESBL-producing genes, typically found on MDR plasmids, which make them resistant to many classes of antimicrobial drugs.19 They can also produce CTX-M-15 cephalosporinases.20 Among all E. coli isolates, ST131 accounts for approximately 12–30% of cases.17 In contrast, its prevalence among fluoroquinolone-resistant and ESBL-producing E. coli isolates is significantly higher, at approximately 70–80% and 50–60%, respectively.17,21
In addition to acquiring MDR genes,22,23 the spread of ST131 has been attributed to its transmission24 and enhanced colonisation abilities.24-26 Although ST131 is not typically associated with hypervirulence,27 it has been observed to outcompete commensal gut E. coli in murine models and is an efficient gut coloniser.28 This literature review will examine how factors such as virulence, MDR gene acquisition, colonisation, and transmission influence the evolution and widespread dissemination of ST131 strains, as well as the interplay among these factors that contributes to their success as a global superbug.
EVOLUTION OF MDR SUB-LINEAGES OF ST131
Recently, the prevalence of MDR E. coli strains has increased.6,17,29 In 2019, MDR E. coli caused just over 800,000 global deaths.6 E. coli is widely classified into phylogroups A, B1, B2, C, D, E, and F, and further categorised into STs.5 ST131 belongs to phylogroup B2 of E. coli, with O25b:H4 as the most common serotype.30 It can be further subdivided into three clades (a monophyletic group) depending on the presence of various FimH alleles, namely H41, H22, and H30 (alternatively named A, B, and C in Petty classification),31,32 and further subdivided into subclades or clones (Figure 1).5,32,33
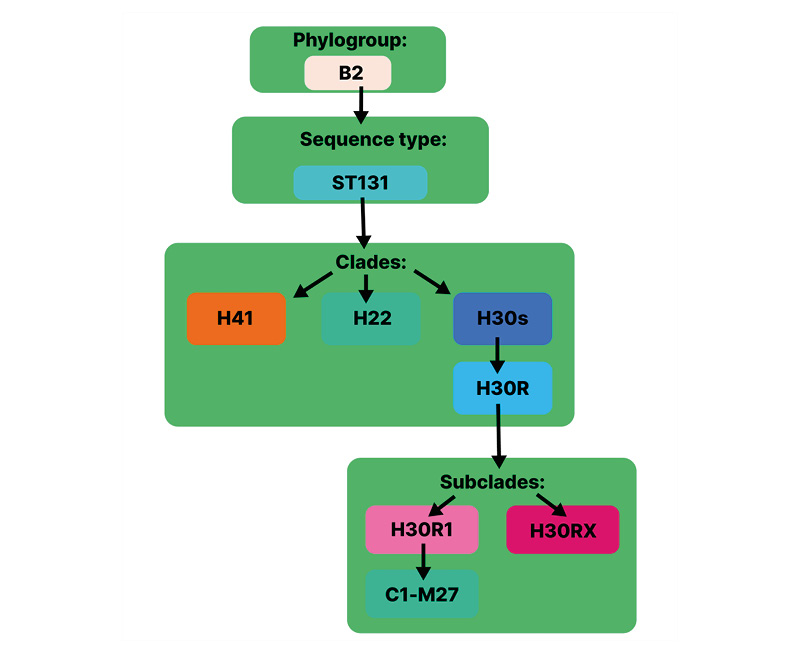
Figure 1: E. coli ST131 division into sub-lineages arranged as phylogroup, sequence type, clades, and subclades.
Adapted from Cummins et al.32
E. coli: Escherichia coli.
ST131 can transmit resistance across species and includes many globally prevalent MDR clones.34 The estimated emergence of its most recent common ancestor is around the mid-1800s.5,35 Resistant ST131-H30R has arisen from drug-susceptible ST131-H30S (alternatively named C0) in 1982.32 The former is divided into fluoroquinolone resistance (FQR) subclades ST131-H30R1 and ST131-H30Rx (Figure 1).32,36 ST131-H30R1 and ST131-H30Rx (containing blaCTX-M-27 and blaCTX-M-15 genes, respectively) are reported to have taken over ST131-H30R (possessing blaCTX-M-14), which was dominant in 2000.37,38 In 2016, subclone C1-M27 emerged in Japan and was soon found to have spread across three continents.5,37 This clade had unique genomic characteristics and was present in ST131 from Thailand, Australia, Canada, and the USA.39 In recent years, the rapidly evolving lineages of these MDR E. coli have been linked to increased international travel and excessive antibiotic use.26,29
Therefore, the rapid emergence and dissemination of lineages containing antibiotic resistance genes can be attributed to multiple factors. Important contributors such as international travel, interspecies transmission, and antibiotic overuse emphasise the urgent need for coordinated stewardship across human, animal, and environmental health.
THE INTERPLAY BETWEEN TRANSMISSION, COLONISATION, AND VIRULENCE, AND ITS INFLUENCE ON THE PROPAGATION OF ST131
The precise reason for the widespread dissemination of ST131 remains unclear; however, effective gut colonisation and its persistence and transmission may contribute to this phenomenon.24 A study conducted in Sweden identified ESBL-producing E. coli among travellers returning from Türkiye, Southeast Asia, India, and North Africa, which are recognised as regions with a high prevalence of this bacterial strain.12 The travellers were tested with rectal swabs collected before travel and around 12–14 days after return. Thirty-two percent of those who initially tested negative for ESBL-producing E. coli and used antibiotics during the trip tested positive upon return. This finding highlights the link between international travel to high-prevalence areas, antibiotic use, and colonisation.12
Moreover, for ExPEC to infect extraintestinal tissue, colonisation of the host intestines is necessary. Therefore, effectively colonising may significantly contribute to its successful spread.40 Studies demonstrate rectal colonisation among asymptomatic individuals.3 In a case study, asymptomatic household members were tested for gut colonisation after two young children experienced a UTI caused by ESBL-producing MDR ST131.24 Over a 19-week period, a total of three faecal samples were collected per person from seven asymptomatic household members, yielding eight different E. coli isolates. The most persistent strain identified was ST131-H30Rx, detected in five household members across all three faecal samples, consistent with the urine isolates from the patients. In contrast, seven other E. coli strains were consistently found in only one or two household members.24 Additionally, a study conducted in Seville, Spain, found that there was an increased risk of rectal colonisation and transmission in both hospital and community settings where there was a higher level of person-to-person contact. This included groups such as elderly patients, catheterised patients, and those who require enhanced basic care or who were using proton pump inhibitors.25 Also, a study found that person-to-person transmission of E. coli ST131 was more prevalent among household members than hospitalised patients, with the carrier state persisting for up to 4 months.41 These findings indicate that increased transmission and colonisation through close contact among asymptomatic individuals and index patients can manifest across all age groups and in both community and hospital settings. This phenomenon contributes significantly to the dissemination of ST131 within various populations.
However, research regarding the virulence capability of ST131 shows conflicting evidence.40 A study in Northwest England found that ST131 did not exhibit hypervirulence compared to other uropathogenic E. coli strains.27 Although the ST131-H30Rx subclone was reported to have a higher virulence score in this study, the effect on clinical outcomes was not significant.27 Additionally, in another study, ST131 in a murine model was not associated with more severe subcutaneous sepsis than non-ST131.42 Similarly, Lavigne et al.43 found that ST131 was less virulent in animal models using Caenorhabditis elegans and zebrafish embryos than non-ST131 strains. Therefore, it is evident from these studies that the prevalence of ST131 may not be attributable to its pathogenic capability (Table 1).21,27,28,42-50 Moreover, Mokady et al.44 reported that virulence factors such as fimH facilitated bacterial colonisation and promoted adaptation to the gut environment rather than causing infection.
Also, more efficient biofilm production has been reported in ST131, possibly enhancing its gut colonisation ability compared to non-ST131 E. coli.45,46 According to Celebi et al.,46 E. coli biofilm production is a significant virulence factor that contributes to antibiotic resistance. Biofilm production by the ST131 strain may shield the bacteria from high antibiotic levels, thereby promoting resistance even at low levels of exposure.46
Another important factor suggested by a few studies is metabolic fitness, which may explain the efficient transmission of ST131 by prolonging colonisation.40,47 Conversely, others found no significant difference in the metabolic fitness of ST131 compared with that of non-ST131.48,49 In one study, ST131 was found to outcompete the commensal E. coli in colonising the intestines of mice.28 Therefore, it is clear that the persistence of ST131 within the host contributes to its virulence rather than enhancing disease causation (Table 1).50
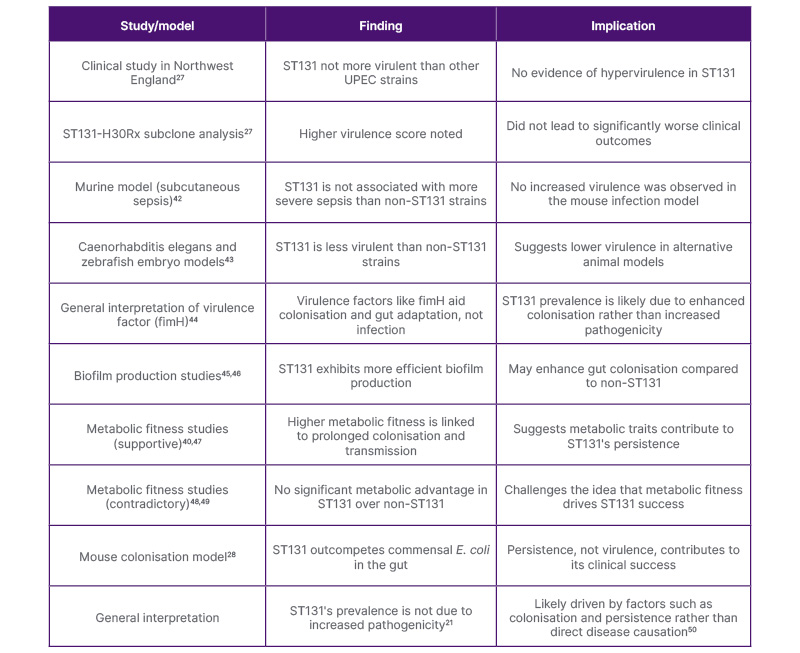
Table 1: Summary of key studies examining the virulence of E. coli ST131, their findings, and implications.21,27,28,42-50
E. coli: Escherichia coli; ST131: sequence type 131; UPEC: uropathogenic E. coli.
THE ROLE OF ANTIBIOTIC RESISTANCE IN ST131 SUCCESS
Numerous studies have demonstrated that antibiotic selection pressure has led to the emergence of successful antimicrobial resistance (AMR) strains.51-53 A common characteristic shared by most ST131 isolates worldwide is resistance to both extended-spectrum cephalosporins and fluoroquinolones.3,16 The production of ESBLs and carbapenemases32 causes resistance to three or more classes of antibiotics,5 including β-lactam antibiotics such as penicillins, cephalosporins, and monobactams.32 Resistance to third generation cephalosporins and carbapenems is associated with the carriage of the CTX-M class of ESBLs and metallo-lactamases, respectively.17,54 H30-Rx has been associated with the spread of ESBL blaCTX-M-15 and ESBL blaCTX-M-14.25 The emergence of clade C has been attributed to the acquisition of FQR through point mutations in DNA gyrase and DNA topoisomerase genes, which are the main drivers, in addition to acquisition of well-defined ExPEC virulence factors.5,33 These factors include adhesins, toxins, iron acquisition factors, lipopolysaccharides, polysaccharide capsules, and invasins, which are usually encoded on pathogenicity islands, plasmids, and other mobile genetic elements.55
Conversely, other studies have shown that the acquisition of MDR was not the main driving force shaping the distribution of ST131, suggesting instead that it was a negative frequency-dependent selection phenomenon (an evolutionary concept where the success of a trait or lineage decreases as it becomes common and increases when rare).5,34 However, many have regarded the ESBL phenotype and specific virulence genes as important factors contributing to their success.10,56,57 Johnson et al.58 observed that the ST131 H30 subclone of E. coli exhibited greater FQR and distinct mutations in gyrA, parC, and parE compared with other fluoroquinolone-resistant strains, suggesting a fitness advantage at low fluoroquinolone concentrations that may aid with their persistence.
AMR genes are located either on chromosomes or plasmids.32 A large MDR plasmid acquired and maintained by ST131 is associated with the carriage of resistance genes40 and their spread within and between the populations.32 The pandemic potential of ST131 is attributed to its MDR ability, which enables it to cause prolonged and more aggressive infections.57 Therefore, the success of E. coli ST131 appears to result from a combination of factors, including multidrug resistance, virulence traits, and evolutionary influences, such as negative frequency-dependent selection (Table 2).3,5,10,16,25,32-34,56-58 While resistance genes are crucial, various other factors also play a significant role in its worldwide dissemination.
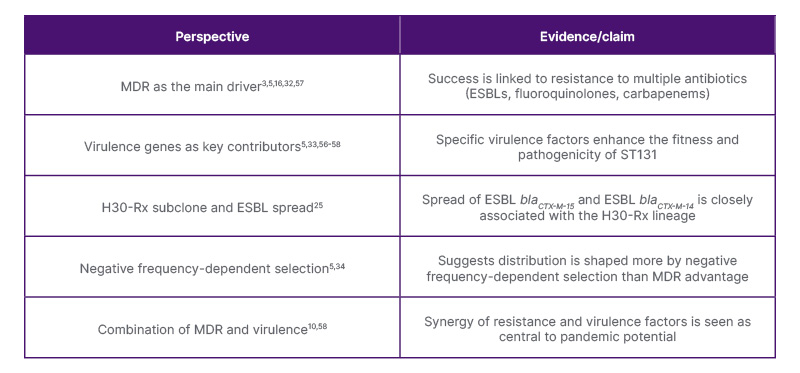
Table 2: Summary of conflicting evidence regarding whether multidrug resistance, virulence, or evolutionary dynamics are the key factors contributing to ST131’s global success.3,5,10,16,25,32-34,56-58
ESBL: extended-spectrum β-lactamase; MDR: multidrug-resistant; ST131: sequence type 131.
IMPORTANCE OF MOBILE GENETIC ELEMENTS IN THE PREVALENCE OF ST131 RESISTANCE
Mobile genetic elements, including plasmids, transposons, bacteriophages, and genetic islands, such as pathogenicity-associated and resistance islands, play a key role in the acquisition of AMR in E. coli ST131.59 Mobile genetic elements encode for the synthesis of virulence factors like siderophores, aerobactin, and yersiniabactin (important for extra-intestinal colonisation), and P fimbrial tip adhesin variant PapGII (PapGII).59,60 PapGII has been associated with higher virulence and the causation of invasive infections through inflammation and renal tissue damage.61,62 A study conducted by Cuénod et al.62 identified PapGII as a major risk factor for progression from UTI to bacteraemia. Similarly, another study found that PapGII was present in isolates from patients with invasive infections, such as bacteraemia or pyelonephritis, whereas these genes were absent from non-invasive (cystitis) or asymptomatic UTI isolates.60
Remarkably, the evolution of ST131 has been transformed by the acquisition of MDR plasmids.50 E. coli ST131 isolates from various origins harbour plasmids that differ in incompatibility group (Inc), conjugative transfer, size, replicon types, and bla genes.3 However, the IncF group of plasmids (related to F-type Pili production) is commonly associated with ST131.63 They carry genes encoding ESBLs21 that contribute to their fitness through determining virulence and AMR.50 Resistance to antibiotics, such as fluoroquinolones, macrolides, and aminoglycosides acquired through IncF plasmids, limits treatment options for infections caused by ST131.63 Additionally, they carry virulence-associated genes, such as: iron haemostasis and enterotoxin production; conjugation genes, for example traT, which encodes a conjugation transfer protein; and toxin-antitoxin systems that maintain plasmids in successive generations of these bacteria.64 In ST131, IncF plasmids utilise addiction systems and post-segregational killing to propagate and maintain themselves, thereby contributing to the spread of blaCTX-M-15 in ST131 isolates, even in the absence of antibiotics.40
Moreover, the acquisition of IncF plasmids led to the emergence of the prevalent H30-Rx subclone.65 Banerjee et al.22 found that H30-Rx isolates had higher virulence scores than other ST131 isolates due to the acquisition of blaCTX-M-15 containing plasmid, which may explain their increased prevalence. Similarly, another study conducted on ST131 prevalence and characteristics among USA veterans found that additional virulence factors aggregated in ST131 as they developed FQR and acquisition of ESBLs, particularly blaCTX-M-15,23 highlighting the role of plasmid-related virulence factors in H30-Rx subclone prevalence. Traditionally, plasmid carriage is associated with a fitness cost, including a decline in bacterial growth rate,50,66 due to plasmid-related cytotoxic effects67 and competition for translational resources between the plasmid and the host cell.68 However, many studies have demonstrated that plasmid carriage did not affect fitness in ST131.69-71 Schaufler et al.71 found that ESBL plasmid-containing strains showed enhanced biofilm formation but reduced motility, as nutrient use was more efficient; the bacteria did not need to move to nutrient-rich areas, resulting in reduced motility. However, in another study, blaCTX-M-14 or blaCTX-M-15 containing ST131 had decreased virulence, particularly in biofilm formation, but no effect on fitness or growth rate compared with the parent ESBL-sensitive strain.69 Therefore, the existence of plasmids in ST131 mutually benefits both50 and may contribute to the success of ST131.
CONCLUSION
The rapid global dissemination of MDR ST131 is a significant public health concern, as numerous frontline antibiotics have become ineffective, particularly in treating common conditions such as UTIs.21 While the specific factors contributing to its global prevalence remain somewhat unclear, evidence indicates that ST131 possesses a notable ability to colonise, persist,24,27 and transmit between hosts,41 even in the absence of traditional hypervirulence traits.47 Its ability to acquire and maintain MDR plasmids has facilitated the horizontal transfer of AMR genes across diverse populations,32 severely constraining treatment options.
To address this escalating threat, a deeper understanding of the host and microbe factors involved in the rapid global dissemination of MDR ST131 is required. Future research should focus on the gut microbiome’s influence on colonisation dynamics and strain competition, as these factors may significantly impact the persistence of ST131. Concurrently, innovative therapeutic approaches targeting colonisation, such as phage therapy and anti-adhesion strategies, could be explored to alleviate transmission. Furthermore, coordinated surveillance through a One Health approach is essential for monitoring the emergence and spread of ST131 across human, animal, and environmental reservoirs. A multidisciplinary effort combining microbiology, clinical research, and public health is critical to developing sustainable strategies to combat this high-risk clone.




